1. Bikbov, B., et al., Global, regional, and national burden of chronic kidney disease, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. The lancet, 2020. 395(10225): p. 709-733. https://doi.org/10.1016/S0140-6736(20)30045-3
2. Chen, J.; Deng, M.; Zheng, R.; Chen, Y.; Pang, W.; Zhang, Z.; Tan, Z.; Bai, Z. Global, regional, and national trends in chronic kidney disease burden (1990–2021): a systematic analysis of the global burden of disease in 2021. Trop. Med. Health 2025, 53(1), 26. https://doi.org/10.1186/s41182-025-00703-x
3. Parsegian, K.; Trivedi, R.; Ioannidou, E. Renal Diseases. Burket's Oral Med. 2021, 579-626. https://doi.org/10.1002/9781119597797.ch16
4. Kalantar-Zadeh, K.; Jafar, T. H.; Nitsch, D.; Neuen, B. L.; Perkovic, V. Chronic kidney disease. Lancet 2021, 398(10302), 786-802. https://doi.org/10.1016/S0140-6736(21)00519-5
5. Vargas, R. A. V.; Millán, J. M. V.; Bonilla, E. F. Renin–angiotensin system: Basic and clinical aspects—A general perspective. Endocrinol. Diabetes Nutr. 2022, 69(1), 52-62. https://doi.org/10.1016/j.endien.2022.01.005
6. Su, C.; Xue, J.; Ye, C.; Chen, A. Role o f the central renin-angiotensin system in hypertension. Int. J. Mol. Med. 2021, 47(6), 95. https://doi.org/10.3892/ijmm.2021.4928
7. Zamani-Garmsiri, F.; Emamgholipour, S.; Rahmani Fard, S.; Ghasempour, G.; Jahangard Ahvazi, R.; Meshkani, R. Polyphenols: potential anti‐inflammatory agents for treatment of metabolic disorders. Phytother. Res. 2022, 36(1), 415-432 https://doi.org/10.1002/ptr.7329
8. Avila-Carrasco, L.; Majano, P.; Sánchez-Toméro, J. A.; Selgas, R.; López-Cabrera, M.; Aguilera, A.; González Mateo, G. Potential therapeutic effects of natural plant compounds in kidney disease. Molecules 2021, 26(20), 6096. https://doi.org/10.3390/molecules26206096
9. Bhullar, S. K.; Dhalla, N. S. Angiotensin II-induced signal transduction mechanisms for cardiac hypertrophy. Cells 2022, 11(21), 3336. https://doi.org/10.3390/cells11213336
10. Rotariu, D.; Babes, E. E.; Tit, D. M.; Moisi, M.; Bustea, C.; Stoicescu, M.; Radu, A. F.; Vesa, C. M.; Behl, T.; Bungau, A. F.; et al. Oxidative stress–Complex pathological issues concerning the hallmark of cardiovascular and metabolic disorders. Biomed. Pharmacother. 2022, 152, 113238. https://doi.org/10.1016/j.biopha.2022.113238
11. Faki, Y.; Er, A. Different chemical structures and physiological/pathological roles of cyclooxygenases. Rambam Maimonides Med. J. 2021, 12(1), e0003. https://doi.org/10.5041/RMMJ.10426
12. Xu, C.; Fang, H.; Zhou, L.; Lu, A.; Yang, T. Na+-retaining action of COX-2 (Cyclooxygenase-2)/EP1 pathway in the collecting duct via activation of intrarenal renin-angiotensin-aldosterone system and epithelial sodium channel. Hypertension 2022, 79(6), 1190-1202. https://doi.org/10.1161/HYPERTENSIONAHA.121.17245
13. Savedchuk, S.; Phachu, D.; Shankar, M.; Sparks, M. A.; Harrison-Bernard, L. M. Targeting glomerular hemodynamics for kidney protection. Adv. Kidney Dis. Health 2023, 30(2), 71-84. https://doi.org/10.1053/j.akdh.2022.12.003
14. Jaimes, E. A.; Tian, R. X.; Pearse, D.; Raij, L. Up-regulation of glomerular COX-2 by angiotensin II: Role of reactive oxygen species. Kidney Int. 2005, 68(5), 2143-2153. https://doi.org/10.1111/j.1523-1755.2005.00670.x
15. Quadri, S. S.; Majid, D. S. Interaction of the renin angiotensin and cox systems in the kidney. Front. Biosci. (Schol Ed) 2016, 8(2), 215-226. https://doi.org/10.2741/s459
16. Zhao, Y.; Liu, Y.; Wang, J.; Ma, X.; Yang, J.; Zhang, L.; Zhang, X.; Yin, J. COX-2 is required to mediate crosstalk of ROS-dependent activation of MAPK/NF-κB signaling with pro-inflammatory response and defense-related NO enhancement during challenge of macrophage-like cell line with Giardia duodenalis. PLoS Negl. Trop. Dis. 2022, 16(4), e0010402. https://doi.org/10.1371/journal.pntd.0010402
17. Aranda-Rivera, A. K.; Cruz-Gregorio, A.; Pedraza-Chaverri, J.; Scholze, A. Nrf2 activation in chronic kidney disease: promises and pitfalls. Antioxidants 2022, 11(6), 1112. https://doi.org/10.3390/antiox11061112
18. Kuczeriszka, M.; Wąsowicz, K. Animal models of hypertension: The status of nitric oxide and oxidative stress and the role of the renal medulla. Nitric Oxide 2022, 125, 40-46. https://doi.org/10.1016/j.niox.2022.06.003
19. Shuvo, A. U. H.; Alimullah, M.; Jahan, I.; Mitu. K. F.; Rahman, M. J.; Akramaddaula, k.; Khan, F.; Dash, P. R.; Subhan, N.; Alam, M. A. Evaluation of Xanthine Oxidase Inhibitors Febuxostat and Allopurinol on Kidney Dysfunction and Histological Damage in Two-Kidney, One-Clip (2K1C) Rats. Scientifica 2025, 2025(1), 7932075. https://doi.org/10.1155/sci5/7932075
20. Gupta, K.; Bagang, K.; Simgh, G.; Arora, S.; Bedi, O.; Kumar, M. Pharmacology of angiotensin in renovascular diseases. In Angiotensin, Elsevier: 2023; pp 151-178. https://doi.org/10.1016/B978-0-323-99618-1.00012-X
21. Okolonkwo, B. B.; Ajibo, D. N.; George-Opuda, I. M.; Donatus, N. J. Recent Trends in Risk Factors Associated with Kidney Diseases. Asian J. Res. Nephrol. 2024, 7(1), 7-30.
22. Wu, R.; Lamontagne, D.; de Champlain, J. Antioxidative properties of acetylsalicylic Acid on vascular tissues from normotensive and spontaneously hypertensive rats. Circulation, 2002. 105(3): p. 387-92. https://doi.org/10.1161/hc0302.102609
23. Wu, R.; Laplante, M. A.; De Champlain, J. Prevention of angiotensin II-induced hypertension, cardiovascular hypertrophy and oxidative stress by acetylsalicylic acid in rats. J. Hypertens. 2004, 22(4), 793-801. https://doi.org/10.1097/00004872-200404000-00023
24. Suleyman, B.; Albayrak, A.; Kurt, N.; Demirci, E.; Gundogdu, C.; Aksoy, M. The effect of etoricoxib on kidney ischemia-reperfusion injury in rats: a biochemical and immunohistochemical assessment. Int. Immunopharmacol. 2014, 23(1), 179-185.https://doi.org/10.1016/j.intimp.2014.06.042
25. Yapca, O. E.; Turan, M. I.; Yilmaz, I.; Salman, S.; Gulapoglu, M.; Suleyman, H. Benefits of the antioxidant and anti-inflammatory activity of etoricoxib in the prevention of ovarian ischemia/reperfusion injury induced experimentally in rats. J. Obstet. Gynaecol. Res. 2014, 40(6), 1674-1679.https://doi.org/10.1111/jog.12373
26. Sultana, S.; Eva, S. A.; Siddiqa, S.; Ghosh, H. C.; Amin, M.S.; Rahman, S; Hossen M.T; Subhan, N.; Alam, M. A. Islam M. N. Black seed (Nigella sativa) powder supplementation prevented oxidative stress and cardiac fibrosis in isoprenaline administered rats. J. Bio. Exp. Pharmacol. 2024, 2(1), 31- 45. https://doi.org/10.62624/JBEP00.00 09
27. Tracey, W. R.; Tse, J.; Carter, G. Lipopolysaccharide-induced changes in plasma nitrite and nitrate concentrations in rats and mice: pharmacological evaluation of nitric oxide synthase inhibitors. J. Pharmacol. Exp. Ther. 1995, 272(3), 1011-1015. https://doi.org/10.1016/S0022-3565(25)24521-9
28. Jahan, I.; Shuvo, A. U. H.; Alimullah, M.; Rahman, A. S. M. N.; Siddiqua, S.; Rafia, S.; Khan, F.; Ahmed, K. S.; Hossain, H.; Akramaddaula, K.; Alam, M. A.; Subhan, N. Purple potato extract modulates fat metabolizing genes expression, prevents oxidative stress, hepatic steatosis, and attenuates high-fat diet-induced obesity in male rats. PLoS One 2025, 20(4), e0318162.https://doi.org/10.1371/journal.pone.0318162
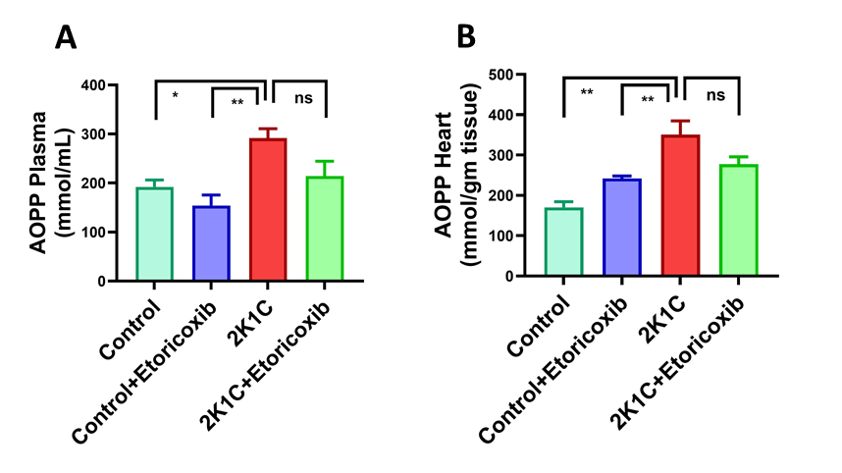
29. Witko-Sarsat, V.; Friedlander, M.; Capeillère-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A. T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49(5), 1304-1313.https://doi.org/10.1038/ki.1996.186
30. Tiwari, B. K.; Kumar, D.; Abidi, A. B.; Rizvi, S. I. Efficacy of Composite Extract from Leaves and Fruits of Medicinal Plants Used in Traditional Diabetic Therapy against Oxidative Stress in Alloxan-Induced Diabetic Rats. ISRN Pharmacol. 2014, 2014, 608590. https://doi.org/10.1155/2014/608590
31. Alimullah, M.; Rahman, N.; Sornaker, P.; Akramuddaula, K.; Sarif, S.; Siddiqua, S.; Mitu, K. F.; Jahan, I.; Khan, F.; Subhan, N.; Alam, A. Evaluation of Terminalia arjuna Bark Powder Supplementation on Isoprenaline-Induced Oxidative Stress and Inflammation in the Heart of Long Evans Rats, Understanding the Molecular Mechanism of This Old Medicinal Plant. J. Med. Nat. Prod. 2024, 100004, 100004–100004. https://doi.org/10.53941/jmnp.2024.100004
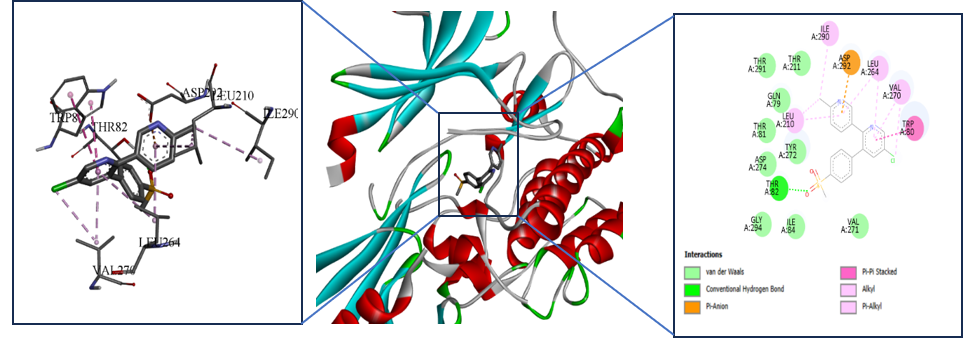
32. Forli, S.; Huey, R.; Pique, M. E.; Sanner, M. F.; Goodsell, D. S.; Olson, A. J. Computational Protein–Ligand Docking and Virtual Drug Screening with the AutoDock Suite. Nat. Protoc. 2016, 11(5), 905–919. https://doi.org/10.1038/nprot.2016.051
33. Trott, O.; Olson, A. J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31(2), 455–461. https://doi.org/10.1002/jcc.21334
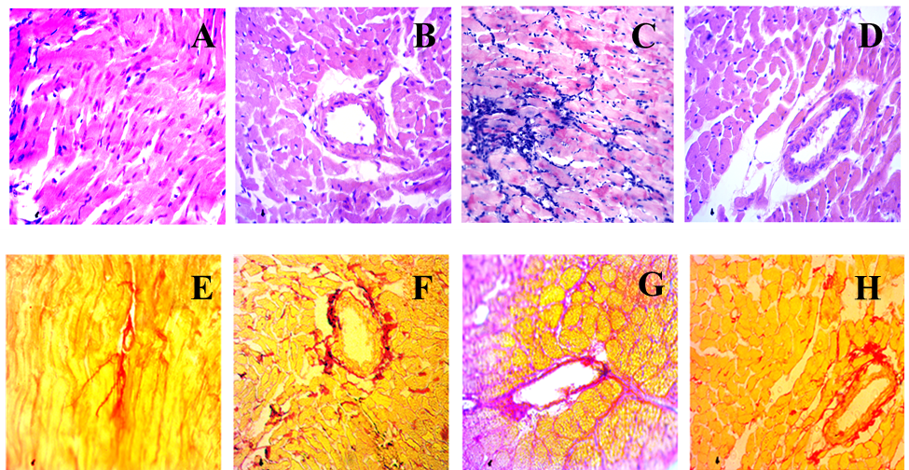
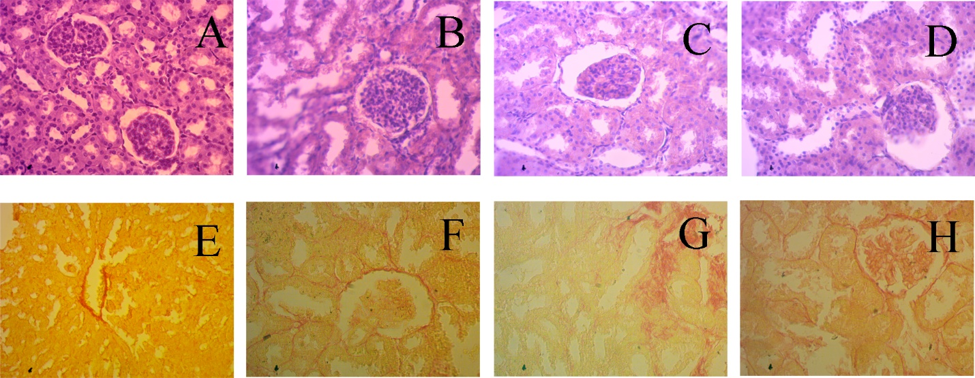
34. de Bhailís, Á. M.; Chrysochou, C.; Kalra, P. A. Inflammation and Oxidative Damage in Ischaemic Renal Disease. Antioxidants 2021, 10(6), 845. https://doi.org/10.3390/antiox10060845
35. Yan, M.-T.; Chao, C.-T.; Lin, S.-H. Chronic Kidney Disease: Strategies to Retard Progression. Int. J. Mol. Sci. 2021, 22(18), 10084. https://doi.org/10.3390/ijms221810084
36. Mansour, A. S. Autoregulation: Mediators and Renin–Angiotensin System in Diseases and Treatments. Future J. Pharm. Sci. 2023, 9(1), 30. https://doi.org/10.1186/s43094-023-00482-4
37. Ghosh, R.; Alajbegovic, A.; Gomes, A. V. NSAIDs and Cardiovascular Diseases: Role of Reactive Oxygen Species. Oxid. Med. Cell. Longev. 2015, 2015(1), 536962. https://doi.org/10.1155/2015/536962
38. Kobiec, T.; Otero-Losada, M.; Chevalier, G.; Udovin, L.; Bordet, S.; Menéndez-Maissonave, C.; Capani, F.; Pérez-Lloret, S. The Renin–Angiotensin System Modulates Dopaminergic Neurotransmission: A New Player on the Scene. Front. Synaptic Neurosci. 2021, 13, 638519. https://doi.org/10.3389/fnsyn.2021.638519
39. Dugbartey, G.J., Gasotransmitters in Organ Transplantation. 2024: Springer. https://doi.org/10.1007/978-3-031-48067-6
40. Zhao, S.; Cheng, C. K.; Zhang, C. L.; Huang, Y. Interplay between Oxidative Stress, Cyclooxygenases, and Prostanoids in Cardiovascular Diseases. Antioxid. Redox Signal. 2021, 34(10), 784–799. https://doi.org/10.1089/ars.2020.8105
41. Leatherby, R. J.; Theodorou, C.; Dhanda, R. Renal Physiology: Blood Flow, Glomerular Filtration and Plasma Clearance. Anaesth. Intensive Care Med. 2021, 22(7), 439–442. https://doi.org/10.1016/j.mpaic.2021.05.003
42. Tsikas, D. Assessment of Lipid Peroxidation by Measuring Malondialdehyde (MDA) and Relatives in Biological Samples: Analytical and Biological Challenges. Anal. Biochem. 2017, 524, 13–30. https://doi.org/10.1016/j.ab.2016.10.021
43. Tirani, M.; Haghjou, M. Reactive Oxygen Species (ROS), Total Antioxidant Capacity (AOC) and Malondialdehyde (MDA) Make a Triangle in Evaluation of Zinc Stress Extension. JAPS J. Anim. Plant Sci. 2019, 29(4).
44. Mazuryk, O.; Gurgul, I.; Oszajca, M.; Polaczek, J.; Kieca, K.; Bieszczad-Żak, E.; Martyka, T.; Stochel, G. Nitric Oxide Signaling and Sensing in Age-Related Diseases. Antioxidants 2024, 13(10), 1213. https://doi.org/10.3390/antiox13101213
45. Pérez de la Lastra, J. M.; Andrés Juan, C.; Plou, F. J.; Pérez-Lebeña, E. The Nitration of Proteins, Lipids and DNA by Peroxynitrite Derivatives—Chemistry Involved and Biological Relevance. Stresses 2022, 2(1), 53–64. https://doi.org/10.3390/stresses2010005
46. Zhou, X.-Y.; Zhang, J.; Li, Y.; Chen, Y.-X.; Wu, X.-M.; Li, X.; Zhang, X.-F.; Ma, L.-Z.; Yang, Y.-Z.; Zheng, K.-M.; Liu, Y.-D.; Wang, Z.; Chen, S.-L. Advanced Oxidation Protein Products Induce G1/G0‐Phase Arrest in Ovarian Granulosa Cells via the ROS‐JNK/p38 MAPK‐p21 Pathway in Premature Ovarian Insufficiency. Oxid. Med. Cell. Longev. 2021, 2021(1), 6634718. https://doi.org/10.1155/2021/6634718
47. Ndrepepa, G. Aspartate Aminotransferase and Cardiovascular Disease—A Narrative Review. J. Lab. Precis. Med. 2021, 6. https://doi.org/10.21037/jlpm-20-93
48. Jewad, A. M.; Jihad, I. A. Role of Heart Failure in Variation of Serum ALT, AST, ALP, Bilirubin and Electrolytes. Biochem. Cell. Arch. 2021, 21(2).
49. Ferrannini, G.; Rosenthal, N.; Hansen, M. K.; Ferrannini, E. Liver Function Markers Predict Cardiovascular and Renal Outcomes in the CANVAS Program. Cardiovasc. Diabetol. 2022, 21(1), 127. https://doi.org/10.1186/s12933-022-01558-w
50. Scholz, H.; Boivin, F. J.; Schmidt-Ott, K. M.; Bachmann, S.; Eckardt, K.-U.; Scholl, U. I.; Persson, P. B. Kidney Physiology and Susceptibility to Acute Kidney Injury: Implications for Renoprotection. Nat. Rev. Nephrol. 2021, 17(5), 335–349. https://doi.org/10.1038/s41581-021-00394-7
51. Moiseyenko, I.O., et al., Summary of European recommendations on nephrology. 2023.
52. Luyckx, V. A.; Rule, A. D.; Tuttle, K. R.; Delanaye, P.; Liapis, H.; Gandjour, A.; Romagnani, P.; Anders, H.-J. Nephron Overload as a Therapeutic Target to Maximize Kidney Lifespan. Nat. Rev. Nephrol. 2022, 18(3), 171–183. https://doi.org/10.1038/s41581-021-00510-7
53. Sethi, S.; De Vriese, A. S.; Fervenza, F. C. Acute Glomerulonephritis. Lancet 2022, 399(10335), 1646–1663. https://doi.org/10.1016/S0140-6736(22)00461-5
54. Anders, H.-J.; Kitching, A. R.; Leung, N.; Romagnani, P. Glomerulonephritis: Immunopathogenesis and Immunotherapy. Nat. Rev. Immunol. 2023, 23(7), 453–471. https://doi.org/10.1038/s41577-022-00816-y
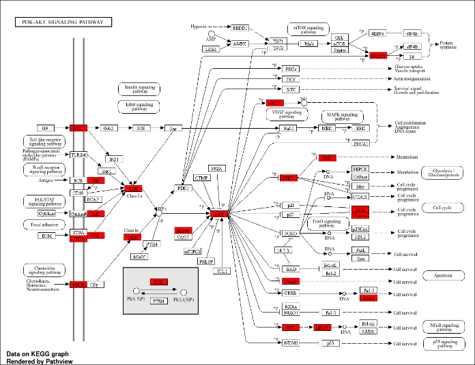
55. He, Y.; Sun, M. M.; Zhang, G. G.; Yang, J.; Chen, K. S.; Xu, W. W.; Li, B. Targeting PI3K/Akt Signal Transduction for Cancer Therapy. Signal Transduct. Target. Ther. 2021, 6(1), 425. https://doi.org/10.1038/s41392-021-00828-5